El homínido de Denísova es el nombre dado a una posible nueva especie de Homo, identificada a través del análisis del ADN de restos óseos encontrados en Siberia en 2010, en la cueva de Denisova. Se piensa que esta especie vivió entre hace un millón y 40 000 años, en áreas en las que también vivían neandertales y Homo sapiens, aunque su origen se encontraría en una migración distinta a las asociadas con humanos modernos y neandertales. En caso de confirmarse como nueva especie de Homo, el nombre oficial podría ser Homo siberiensis, Homo altaiensis o más probablemente Homo denisoviensis.
Un equipo de científicos del Instituto Max Planck de Antropología Evolutiva en Leipzig, Alemania, liderado por Svante Pääbo, secuenció ADN mitocondrial extraído de un fragmento de hueso proveniente del dedo de una niña que fue encontrada en las cuevas de Denísova. Este estudio ha detectado evidencia genética que apunta a su hibridación con poblaciones humanas modernas, pero solo con poblaciones indígenas de Australia, Nueva Guinea y áreas circundantes. Por el contrario, el ADN del homínido de Denisova parece estar ausente o en niveles muy bajos en las poblaciones actuales en el continente asiático, a pesar de que éste es el lugar donde se encontró el fósil.
Un molar descubierto en la misma cueva también proporcionó ADN mitocondrial que se asemeja al del hueso del dedo. El molar presenta características morfológicas claramente diferentes a las de los neandertales y los humanos modernos, lo que confirma que pertenece a una especie con una historia evolutiva distinta.
Por otra parte, la secuenciación de ADN mitocondrial de un fémur procedente del yacimiento de la Sima de Los Huesos, en Atapuerca, de hace 400.000 años, ha mostrado mayor relación con el ADN denisovano que con el neandertal, lo que abre paso a la hipótesis de una compleja relación entre las distintas especies de Homo en Eurasia.
Una vez establecida como especie diferente de neandertales y H. sapiens, la investigación se dirige hacia establecer la relación con otros fósiles encontrados en Asia, en especial los de Homo erectus encontrados en Java y China y los de otros homínidos, como el cráneo de Dali (China), fechado hace 200 000 años, la mujer de Jinniushan, la mandíbula de Zhirendong de hace 106 000 años, el cráneo de Xuchang de 100 000 años y del hombre de Solo (Java), cuyas características morfológicas tienen diferencias y coincidencias con las de H. erectus y H. sapiens. Por otra parte, todo parece indicar que la especie de los denisovanos se dispersó ampliamente en el pasado, y además, que los actuales humanos de Papúa, Nueva Guinea, Australia y Melanesia tienen entre un 4% y un 6% de material genético de los homínidos de Denísova, así como los europeos tienen un 2,5% del genoma heredado de los neandertales.
En una investigación, cuyos resultados se publicaron a finales del 2013, el ADN mitocondrial de la Sima de los Huesos, en España, es muy próximo al de la población representada por los homínidos de Denisova, que vivieron 300.000 años más tarde y a 8.000 Km de distancia. Otra conclusión de esta investigación es que la separación de linajes entre humanos neandertales y humanos completamente modernos fue muy posterior a la separación de los denisovianos respecto a ambos.
Homo erectus es un homínido extinguido, que vivió entre 300.000 años y 1,8 millones de años, en el Pleistoceno inferior y medio.
El hombre de Java (Homo erectus erectus), fue el primer representante de Homo erectus en ser descubierto. Fue encontrado y descrito originalmente en Java por el científico, de profesión médico, Eugène Dubois quien lo nombró inicialmente Pithecanthropus erectus. La palabra pithecantropus deriva de raíces griegas y significa hombre mono.
Dubois encontró los restos en el lugar de Trinil en la Isla de Java en 1891. El hallazgo consistió en la tapa de un cráneo. Un año después fueron descubiertos un fémur y dos muelas a dieciséis metros de donde se encontró la tapa de la calavera. Dubois consideró que todas las piezas provenían del mismo individuo y las fechó con una antigüedad de medio millón de años. Además, calculó la capacidad craneana en 855 cm³.
En la década de 1930 el paleontólogo alemán Ralpf von Koenigswald obtuvo nuevos fósiles, tanto cerca de Trinil como en nuevas localidades como Sangiran (a unos 75 km). En 1939 von Koenigswald y Franz Wiedenreich trabajaron para definir una relación precisa entre los fósiles de Trinill y Sangiran con los del Hombre de Pekín denominado por entonces Sinanthropus pekinensis y concluyeron destacando las semejanzas claves entre ellos que demuestran que representan una misma forma humana antigua. En 1940 Weidenreich reinterpretó los restos de Trinil y Java como Homo erectus iavanensis, pero se renombraron definitivamente por Dobzhansky (1944) como Homo erectus erectus.
Restos de Homo erectus descubiertos por Dubois
Dubois nunca aceptó que los fósiles que encontró en Trinil pudieran relacionarse como una misma especie o forma humana con los de Trinil y Sangiran. También Dubois había encontrado dos calaveras obviamente humanas a 100 km en Wadjak un yacimiento de fecha dudosa, que podía datar de hace 50.000 años. Los detractores de la evolución acusaron a Dubois ocultar esta información para dar más credibilidad a sus otros descubrimientos. Nuevas pruebas sobre la fauna del mismo estrato marcan que estos cráneos, llamados Wadjak 1 y 2, son del Holoceno, lo que los desvincula completamente del Homo erectus.
Homo erectus vivió en Asia oriental (China, Indonesia). En África se han hallado restos de fósiles afines que con frecuencia han sido incluidos en otra especie, Homo ergaster. En Europa, los restos fósiles hallados en Italia y España, también han sido clasificados como Homo erectus, pero muchos especialistas piensan que es acertado, en principio, reservar el nombre Homo erectus para los fósiles asiáticos.
Uno de los rasgos más comúnmente asociados con el Homo erectus es el aumento del tamaño del cuerpo. Poseía un cuerpo robusto con una media de 1,70-1,80 metros de altura, pesaba cerca de 70 kilogramos y caminaba en posición erecta. Posiblemente se desplazaba en pequeños grupos familiares, vivía en cavernas y cazaba en los bosques.
Tenía una capacidad cerebral mayor que la del Homo habilis. Los primeros restos que se encontraron del Hombre de Java muestran una capacidad craneal de 850 centímetros cúbicos, superior a los 650 cc del Homo Habilis. mientras que los que se encontraron posteriormente llegan a los 1.100 cc. Su capacidad craneal media, de unos 940 cc, es intermedia entre los 1.200-1.500 del hombre moderno y los 600 cc del gorila. El cerebro del hombre de Java era mucho más grande y con un mayor número de circunvoluciones que el del cualquiera de los monos primitivos o vivientes, y tenía más características humanas que simiescas.
A medida que el tamaño del cuerpo de los homínidos aumenta, también lo hace el tamaño del cerebro. Además del aumento absoluto en el volumen cerebral que acompaña a un aumento en el tamaño del cuerpo, también hay un aumento proporcional. Esto se conoce como encefalización, y es una característica importante de Homo erectus. A lo largo de la historia evolutiva de Homo erectus hay pruebas sustanciales de selección dirigida a aumentar la encefalización, de modo que mientras los primeros miembros del linaje tienen una capacidad craneal de 600 a 800 cc, las capacidades craneales de la mayoría de los ejemplares posteriores están muy por encima de 1.000 cc, que está dentro el rango inferior de los humanos contemporáneos, sin que parezca mucho más grande en tamaño corporal que los principios de Homo erectus.
El gran cuerpo y cerebro grande del Homo erectus necesita más energía, y por tanto más alimentos que los homínidos anteriores. Estructuras biológicas más grandes, sobre todo las de alto consumo energético, como los músculos y cerebros, requieren mayores consumos de energía para mantenerlos. Por lo tanto, Homo erectus a menudo es reconstruido como ocupando un nicho ecológico intensificado. El nicho intensificado va de la mano con la expansión del cerebro y el tamaño corporal. Cuerpos más grandes y extremidades más largas aumentan la eficiencia del aparato locomotor.
Homo erectus podría cubrir más terreno de día a día, a través de caminar o correr, que los homínidos más pequeños y con menor coste energético. Además, el cerebro más grande dio estos homínidos mejores capacidades para el procesamiento de información ecológica compleja a través del terreno, proporcionándole una alimentación de mayor calidad. Hay una evidencia clara de que Homo erectus comió cadáveres de animales de mediano y gran tamaño a través de la caza o utilización de sus restos, ya que se han encontrado restos fósiles de animales con marcas de corte que indican una comida carnívora. Este comportamiento, carnívoro, es un cambio ecológico respecto a los homínidos anteriores. Mientras que los primeros especímenes de Homo erectus se encuentran en asociación con las herramientas de piedra muy básicas, que se refieren típicamente como la industria lítica olduvayense, hace 1.500.000 años las poblaciones de Homo erectus fueron creando un conjunto más complejo de herramientas conocidas como la industria lítica achelense.
Poseía un marcado toro supraorbitario y una fuerte mandíbula sin mentón, pero de dientes relativamente pequeños.
El cambio en la ecología asociada a Homo erectus coincide con un cambio correspondiente en la dentición y las mandíbulas de esta especie. En relación a los australopitecinos, el tamaño de la dentición de premolares y molares se reduce drásticamente en Homo erectus. El cuerpo de la mandíbula (es decir, la parte sin dientes que se une al cráneo) también muestra un aumento de gracilidad o esbeltez, con una reducción característica en la amplitud relativa de la estructura y de apoyo de las estructuras de masticación. Parece ser que Homo erectus tenía una dieta diversa y amplia. La comida que ingiere un organismo puede producir cambios sutiles en la química de los tejidos del cuerpo, incluyendo la dentina y el esmalte que forman la corona de un diente. A partir de esta información, las investigaciones realizadas a través de la química de isótopos estables de Homo erectus, también apoyan la idea de una dieta flexible y diversa. Cualquiera que sea la flora y la fauna, Homo erectus comió en las diversas zonas geográficas donde se encuentran sus fósiles. Sus dientes y la anatomía de la mandíbula revelan que su dieta no requería las mismas adaptaciones masticatorias robustas, como las de los australopitecus.
Reconstrucción de Homo erectus
Es objeto de discusión científica si presentaba un mayor dimorfismo sexual que en el hombre moderno. El dimorfismo sexual, las diferencias físicas entre hombres y mujeres, es una fuente importante de variación dentro de las especies, y en los primates puede ser un indicador de la estrategia reproductiva y la dinámica de grupo. El dimorfismo sexual, dado su papel en la variación intraespecífica, también puede ser un factor de confusión en la identificación taxonómica. La gran cantidad de variación en el tamaño observado en Homo erectus, tomados principalmente de restos fósiles fragmentarios, hace difícil estimar los niveles medios de dimorfismo. El registro fósil de Homo erectus proporciona una evidencia clara de una amplia gama de variación en el tamaño del esqueleto, que como mínimo, equivale a la observada en las poblaciones humanas que viven, pero no proporciona pruebas concluyentes de que los hombres fueron sistemáticamente mayores que las hembras, en mayor medida de lo que son hoy. Si Homo erectus tenía dimorfismo más sexual que Homo sapiens, es fácil concluir que la competencia masculina dependía más del tamaño del cuerpo que hoy en día.
El hombre de Java poseía la porción del cerebro que controla el lenguaje, aunque se ignora si efectivamente hablaba. El lenguaje es tal vez el rasgo humano distintivo, pero puede ser difícil de evaluar directamente desde el registro fósil. Los intentos de identificar la capacidad lingüística en los restos óseos fosilizados de Homo erectus se han centrado en los aspectos del sistema nervioso, incluyendo el tamaño del canal vertebral, que es una aproximación para el tamaño de la médula espinal, y las características externas de endomoldes (fósiles naturales del espacio endocraneal y estimar el tamaño del cerebro y la forma). Hasta el momento, no ha habido hallazgos anatómicos definitivos, por lo que los investigadores rechazan la idea de que el Homo erectus era capaz de hablar algo similar a una idioma primitivo.
Más recientemente, los argumentos acerca de los orígenes del lenguaje se han centrado en las historias reconstruidas de los genes asociados con la producción del lenguaje en los seres humanos. La recuperación de las antiguas secuencias genéticas de los neandertales y otros especímenes humanos arcaicos (por ejemplo, una muestra de la Cueva Denisova en Siberia, Rusia) han proporcionado una nueva visión de la historia genética de la producción del lenguaje. Los estudios genéticos de genomas asociados con el lenguaje sugieren que este gen probablemente se remonta al menos al Pleistoceno Medio. No existe ADN de Homo erectus para realizar la investigación, pero todo parece indicar que el lenguaje está claro sólo con neandertales y Homo sapiens, aunque tampoco existen evidencias para rechazar la idea de que Homo erectus no poseyera capacidades lingüisticas.
Probablemente dominaba el fuego, ya que la cueva de Black, próxima a Pekín, contenía restos de hogueras, lo cual significa que había sido descubierto el fuego hace unos 500 000 años.
En definitiva, Homo erectus representa una importante transformación de los homínidos anteriores, como los australopitecos, a una especie mucho más parecida a los humanos modernos. Era más grande, más inteligente y más capacitado para ocupar y sobrevivir en diferentes paisajes en un mundo cambiante.
Australopithecus afarensis, homínido australopecino, vivió hace entre los 3 y 3,7 millones de años. Era de morfología delgada y grácil. La mayoría de la comunidad científica considera que puede ser un antepasado del género Homo.
Se piensa que solo vivió en una zona concreta, en el este de África (Etiopía, Tanzania y Kenia).
Fue descubierto el 24 de noviembre de 1974 por Donald Johanson, Yves Coppens y Tim White en el yacimiento de Hadar, en el valle del río Awash, en Etiopía. El espécimen encontrado en aquel momento fue mundialmente conocido como Lucy, que viene de la canción Lucy in the sky with diamonds del grupo musical The Beatles, que escuchaban los miembros del grupo investigador la noche posterior al hallazgo.
Lucy era el Australopithecus mejor conservado hasta entonces. Los restos de Lucy fueron encontrados en el lugar donde habitaba la tribu Afar, de ahí el nombre Australopithecus afarensis. Junto a este, había otros doce individuos de la misma especie, incluido cráneos.
Reconstrucción de Lucy en el Museo de Historia Natural de Cleveland
Su descubrimiento hizo ver que la capacidad para caminar erguido, en posición bípeda, como los humanos actuales, fue muy anterior al desarrollo del cerebro. El estudio de su dentadura aclaró aspectos fundamentales sobre la evolución de los homínidos y descubrió la evolución simultánea de géneros, de esta manera manera que la línea de los homínidos Paranthropus, se separó de otras y en particular de la que evolucionó hacia Homo.
Australopithecus afarensis es, como todos los australopitecinos, un primate bípedo erguido, pero diferente a los humanos en varios aspectos. El dedo gordo del pie se dirige a la línea media, y ya no es prensil, y por tanto sin capacidad de agarrar con el mismo (una característica de los otros primates). El pie modificado de Australopithecus afarensis muestra que los australopitecinos poseen un pie modificado para conseguir una mayor eficiencia del aparato locomotor bípedo, a expensas de las capacidades de escalada. Probablemente era bastante un bípedo caminante eficiente en distancias cortas, y la separación de las huellas en Laetoli indica que iban caminando a 1 m/s, o por encima, lo que coincide con las pequeñas ciudades la velocidad de marcha humanos. Sin embargo, esto puede ser cuestionado, ya que las huellas de Laetoli puede que no correspondan al Australopithecus. Muchos científicos también dudan del bipedismo, y argumentan que incluso si el Australopithecus caminó con dos piernas, no anduvo de la misma manera que los seres humanos. Algunos estudios sostienen que su vida era en parte arbórea. La anatomía de las manos, los pies y las articulaciones de los hombros favorece esta última interpretación. En particular, la morfología del omóplato parece ser simiesca y muy diferente de los humanos modernos.
Las falanges de los dedos eran curvadas, tanto de las manos como de los pies, de forma más marcada en estos últimos. Esta característica sugiere que tenían gran capacidad de subir y colgarse de las ramas de los arboles, por lo que se piensa que los arboles eran su medio más o menos habitual.
Los cambios climáticos que ocurrieron hace unos 10-11 millones de años, afectaron a los bosques de África central y oriental, ya que los claros y zonas despejadas en los mismos, ya no permitían viajar a través de la copa de los árboles, y en estos tiempos los homínidos ancestrales podrían haberse adaptado al comportamiento de caminar en posición vertical para andar por el suelo, mientras que los antepasados de los gorilas y los chimpancés se volvieron más especializados en la vida arbórea, con posturas de cadera doblada, rodilla doblada, y caminata de nudillos en el suelo. Este australopecino habría hecho uso del bipedismo en posición vertical para moverse por la tierra, sin dejar de tener los brazos bien adaptadas para trepar a los árboles más pequeños. Sin embargo, los chimpancés y los gorilas, que son los parientes vivos más cercanos a los seres humanos, comparten características anatómicas con Australophitecus afarensis, incluyendo un hueso de la muñeca fusionado, mecanismo de bloqueo de la misma, que también podría indicar la caminata de nudillos por los antepasados humanos.
La articulación del hombro también está orientada mucho más cranealmente (hacia el cráneo) que en humanos modernos, pero similar a la de los actuales simios. En combinación con los brazos relativamente largos, se piensa que Australopithecus afarensis ha tenido una mayor capacidad para usar el brazo por encima de la cabeza en la escalada. Por otra parte, los análisis de los cráneos revelan una morfología ósea, que no apoya una locomoción bípeda completa.
Su capacidad craneal era bastante menor que la humana, 375 a 550 centímetros cúbicos. Tenía un cerebro del tamaño del de un chimpancé y cercano a la tercera parte del humano actual promedio, aunque en comparación con el tamaño del cuerpo era relativamente grande. Además, su cara era bastante grande y se proyectaba delante del cráneo, debido al tamaño de los dientes (prognatismo). El cráneo presenta pequeñas crestas sagitales y nucales, comparables a los del gorila macho actual, pero mucho más pequeñas.
A diferencia de otros homínidos, los caninos de Australopithecus afarensis son reducidos aunque se proyectan ligeramente delante del diente adyacente. Los incisivos son grandes, asociados al régimen frugívoro. Los molares y premolares son de notable tamaño, con superficies planas. De su boca también se concluye que el paladar es muy similar al humano actual porque aunque es grande, forma una curva que no es parabólica, ni de lados paralelos, como en los grandes simios.
Su pecho no tenía forma de barril, como en los humanos, sino que se estrechaba agudamente hacia arriba (forma de campana).
El dimorfismo sexual era muy marcado, siendo los machos mucho más corpulentos que las hembras, con un promedio de masa corporal de 45 kg para los machos y 29 kg para las hembras.
Reconstrucción de Lucy en la Institución Smithsoniana
La pelvis es mucho más similar a la humana que a la de los simios. Las palas ilíacas son cortas y anchas, el sacro es amplio y se coloca directamente detrás de la articulación de la cadera. La evidencia de un fuerte apego a los extensores de la rodilla es clara.
La pérdida de la posibilidad de agarre de la extremidad posterior aumentaría el riesgo de caída de una cría, al contrario de lo que ocurre, por ejemplo, en orangutanes y monos arañas. La cría no hubiera podido mantener un agarre tan fuerte, sin la ayuda de la madre. El problema de agarrar a la cría se multiplica si la madre también tiene que subir a los árboles.
Los dientes, la mandíbula y la configuración del cráneo indican el tipo de alimentación consumida.
Una parte importante de esta distinción es en el aparato masticatorio. Así, los australopitecinos gráciles tienen una gran dentadura y un gran aparato masticatorio, pero es todavía más grande en algunos de los australopitecos robustos posteriores, en los que estas estructuras se vuelven masivamente robustas. Y en la transición hacia Homo, empezamos a ver como algunas de estas estructuras se reducen drásticamente de forma continua en tamaño.
Cráneo de Australopithecus aethiopicus
Mandíbula de Australophitecus afarensis
Cráneo de Paranthropus conocido como "cascanueces"
Como se puede observar en las anteriores fotografías, el Australophitecus tiene premolares más pequeños que los molares, mientras el Paranthropus "Cascanueces" descubierto por Mary Leakey tiene premolares grandes, estos se expanden considerablemente en relación con el tamaño de los molares.
Resaltar las diferencias entre el tamaño de los premolares de los autralopitecinos gráciles y los robustos. Establecer una diferencia cuantitativa, aunque sea de un modo aproximado. Asimismo indicar que tipo de alimentación estaría asociado con cada dentición.
Los cambios asociados con la masticación no se limitan a los dientes. También existen diferencias registradas en las estructuras de la mandíbula y el cráneo asociado con los músculos más utilizados en la masticación. Así, el tamaño de los arcos cigomáticos indica la potencia de los músculos maseteros. Una de las diferencias más notable es la robustez de la sección transversal de estas mandíbulas. Las mandíbulas robustas suelen ser muy amplias en relación con la altura.
Indicar las diferencias, en el sentido del tamaño de los arcos cigomáticos y de la forma y robustez de las mandíbulas, de las especies de homínidos de la imagen superior.
2. El foramen magno
El foramen magno o agujero magno (foramen magnum) es el nombre del orificio situado en la parte inferior del cráneo, en su base, a través del cual se establece la continuidad del sistema nervioso. Por él pasa la médula espinal, que da sensibilidad a todo el resto del cuerpo.
Una de las grandes diferencias anatómicas de la especie humana con los demás homínidos es el ángulo de la articulación del atlas con el foramen magnum. En los homínidos el atlas articula en un plano oblicuo con el foramen magnum, mientras que en el hombre el plano de esta articulación es paralelo al suelo. En la evolución del hombre, cuanto más abajo estuviera el foramen magno, más erguida era la posición de la especie, lo que implicaba un mayor bipedismo.
En el Australopithecus afarensis, el foramen magno aparece en una posición intermedia, un poco más hacia atrás que en el hombre, pero no tanto como en el chimpancé.
Foramen magno en gorila y humanos
En las imágenes inferiores aparece mostrado la posición del foramen magno en el chimpance, en el Australopithecus y en la especie humana. Poner de relieve las diferencias, mostrando su relación con el bipedismo.
Foramen magno en chimpance, Australopithecus y especie humana
La gran visión de Darwin y el principio unificador de la biología actual es que todas las especies se relacionan entre sí como hermanas, primas y parientes lejanos en el gran árbol de la vida. Las implicaciones son impresionantes, y si pudiéramos viajar atrás lo suficientemente lejos en el tiempo, nos encontraríamos con antepasados comunes entre nosotros y cualquier otro organismo vivo, del puerco espín de los flamencos a los cactus. Nuestra familia evolutiva inmediata se compone de los homínidos , el grupo de primates que incluye a los "simios menores" (siamangs y gibones), así como los "grandes simios" (chimpancés, bonobos, gorilas y orangutanes). Entre los grandes simios, nuestros parientes más cercanos son los chimpancés y los bonobos. El registro fósil, junto con los estudios de ADN humano y simio, indican que los seres humanos comparten un ancestro común con los chimpancés y los bonobos en algún momento alrededor de hace 6 millones de años. La divergencia de nuestro linaje con chimpances y bonobos comienza en África, en algún momento al final del período de tiempo geológico conocido como el Mioceno.
Los orígenes del linaje homínido en el Mioceno
Para comprender la evolución de las especies, debemos primero establecer su estado ancestral: ¿qué tipo de animal es el que evolución hacia los homínidos? Para ello debemos reconstruir el último ancestro común de los seres humanos y los chimpancés y bonobos y que a partir de él divergieron. Esta nuestra rama del árbol del linaje de los homínidos incluye todas las especies extinguidas y la evolución de ramas laterales.
Existía una gran diversidad de especies de monos en el Mioceno, con decenas de especies conocidas en el registro fósil a través en África, Europa y Asia. Estas especies eran variadas en su anatomía y su ecología, y ninguna de las especies fósiles descubiertos hasta el momento representan el último ancestro común entre humanos y chimpancés. Sin embargo, sabemos por la evidencia fósil que era mucho más parecido a los simios actuales que a los seres humanos vivos. Habría tenido un cerebro y un cuerpo del tamaño de un mono, con brazos y dedos relativamente largos y un pie con agarre que permitiría andar por los árboles. Los dientes caninos eran probablemente grandes y fuertes, como se ha visto en varios homínidos del Mioceno. Por otra parte, los caninos fueron probablemente un componente del dimorfismo sexual, en el que los machos tienen colmillos mucho más grandes que las hembras, como se ha visto entre los grandes simios vivos y fósiles del Mioceno. Al igual que los monos que viven ahora, habría caminado de forma cuadrúpeda (en cuatro patas) estando en tierra, y su dieta habría consistido casi enteramente de alimentos de origen vegetal, principalmente de fruta y hojas.
Los primeros homínidos
Los cambios de una anatomía parecida al simio son discernibles en los homínidos fósiles del Mioceno tardío en África. Algunas especies de homínidos de esta época muestran características típicas de los humanos, que no se ven en los otros simios vivientes, lo que lleva a los paleoantropólogos a inferir que estos fósiles representan los primeros miembros del linaje homínido. Los primeros rasgos parecidos a los humanos que aparecen en el registro fósil de los homínidos son el caminar bípedo y el menor tamaño de los caninos .
Los homínidos más antiguos conocidos actualmente son Sahelanthropus tchadensis de Chad y Orrorin tugenensis de Kenia. Sahelanthropus tchadensis vivió hace 6-7 millones de años. Se conoce a partir de un cráneo en gran medida completo y algunos otros restos fragmentarios. Su tamaño del cerebro, 360 cm³, está dentro del intervalo observado en los chimpancés, y el cráneo tiene un arco superciliar prominente, con un grosor similar al de los gorilas machos. Sin embargo, la posición y la orientación del foramen magno , el orificio de la base del cráneo a través del cual pasa la médula espinal, sugiere que Sahelanthropus se puso de pie y caminó bipedamente, con su columna vertebral en posición vertical como en los humanos modernos en lugar de horizontalmente, como en los simios y otros cuadrúpedos. Orrorin es conocido principalmente por restos fósiles postcraneales, incluyendo partes de fémur. La parte próxima del fémur muestra similitudes con los de los humanos modernos, lo que sugiere la especie era bípeda. No se han descubierto cráneos de Orrorin, por lo que de su morfología craneal y del tamaño de su cerebro no hay seguridad. En tanto que en Orrorin y Sahelanthropus los dientes caninos de los machos son más grandes y más acentuados que en los seres humanos modernos, pero son pequeños y romos en comparación con los caninos de monos macho, esto sugiere que existía un dimorfismo sexual en los caninos y, por extensión, competencia entre machos por el apareamiento con las hembras, que se redujo en estos primeros homínidos en comparación con los grandes simios.
El mejor homínido temprano conocido es el Ardipithecus ramidus, una especie de hace 4,4 millones de años en Etiopía, que se conoce a partir de un esqueleto casi completo, así como otros numerosos restos dentales y esqueléticos. Esta especie se ha relacionado con Ardipithecus kadabba, con una edad de 5,2-5,8 millones de años, en la que se han reducido los caninos, similares a los de Orrorin y Sahelanthropus. El cráneo de Ardiphitecus ramidus es bastante parecido al de un mono y muy similar al de Sahelanthropus, con un cerebro pequeño como de chimpancé de tamaño de 300-350 cm³. El esqueleto postcraneal de Ardipithecus es intrigante. Aunque muy fragmentación, la pelvis recuperada revela una morfología bastante diferente de la de los simios actuales, con una forma más corta, más parecida a un tazón que sugiere fuertemente que los Ardipithecus caminaron a dos patas, lo que es coherente con la posición del foramen magnum, sugiriendo una postura erguida. Sin embargo, sus largas extremidades y dedos, y la posición del agarre del primer dedo del pie (hallux) sugieren que Ardipithecus pasó gran parte de su tiempo en los árboles. La impresión general es de una especie arbórea en gran medida, pero que caminaba bipedamente cada vez que se aventuró en el suelo.
Australopithecus
Alrededor de hace 4 millones de años aparecen los primeros miembros del género Australopithecus, homínidos bípedos terrestres pero que no obstante seguían utilizando los árboles para alimento y protección. Los primeros ejemplares de Australopithecus fueron descubiertos en Sudáfrica en 1924, y los esfuerzos de investigación en las siguientes ocho décadas han fructificado en el descubrimiento de cientos de fósiles de varias especies en todo África oriental y meridional. Ahora sabemos que el Australopithecus era un género de gran éxito que se mantuvo durante casi tres millones de años.
Los más conocidos Australopithecus son las especies Australopithecus afarensis (2,9-3,6 millones de años) en el este de África y Australopithecus africanus (2,0-3,2 millones de años) de Sudáfrica. La pelvis y las extremidades inferiores de estas especies indica claramente que eran totalmente bípedos, la pelvis es corta y en forma de cuenco, con lo que los músculos de los glúteos, al igual que en los seres humanos modernos, sirven para la estabilización del tronco durante la bipedestación, y el primer dedo del pie está en línea con los otros dedos. El pie del Australopithecus puede incluso haber tenido una morfología similar a la humana, es base al análisis de los metatarsianos y las huellas de Laetoli fosilizadas. Sin embargo, en comparación con los humanos modernos, los brazos eran largos y los dedos eran largos y algo curvados, lo que sugiere que el Australopithecus utilizaba regularmente los árboles para alimentarse y tal vez como un refugio de los depredadores durante la noche. Esta estrategia mixta terrestre y arbórea mixta habría utilizada también en los ambientes forestales y de sabanas mixtas que habitaban.
El tamaño del cerebro en Australopithecus osciló entre 390 y 515 centímetro cúbicos, similar a los chimpancés y gorilas, lo que sugiere habilidades similares a las de los simios vivos. El tamaño del cuerpo de Australopithecus era bastante pequeño y presentaba dimorfismo sexual, alrededor de 30 kg para las hembras y de 40 kg para los machos. Este nivel de dimorfismo no se refleja en los caninos, que eran pequeños, contundentes y monomórficos como en los homínidos anteriores.
A diferencia de los caninos, los dientes molares en Australopithecus eran mucho más grandes que los de los homínidos anteriores, y tenían un esmalte más grueso. Esto sugiere que su dieta incluía alimentos de origen vegetal, de baja calidad, difíciles de masticar. Un subgrupo de Australopithecus, conocido como los australopitecinos "robustos" (representado por un género separado, Paranthropus), debido a sus enormes dientes y los músculos masticadores, llevaron esta adaptación al extremo. La mayoría de las especies de Australopithecus se habrían extinguido hace 2 millones de años, pero algunas formas persistieron hasta cerca de 1,2 millones de años en África Oriental y del Sur.
El género Homo
Los fósiles más antiguos de nuestro propio género, Homo, se encuentran en el este de África con fecha de 2,3 millones de años. Estos primeros ejemplares son similares en el cerebro y el tamaño corporal de Australopithecus, pero muestran diferencias en sus molares, lo que sugiere un cambio en la dieta. En efecto, hace por lo menos 1,8 millones de años, los primeros miembros de nuestro género utilizaban herramientas primitivas de piedra para descuartizar cadáveres de animales, agregando energía, carne y médula ósea a su dieta a base de plantas.
El miembro más antiguo del género Homo, Homo habilis (1,4-2,3 millones de años) aparece en el este de África y está asociado con huesos de cadáveres animales y herramientas de piedra simples. Su descendiente más formidable y exitoso, Homo erectus, se encuentra en toda África y Eurasia, y se mantuvo entre 100.000 a 1,9 millones de años, y quizás incluso más tarde. Al igual que los seres humanos modernos, Homo erectus carecía de las adaptaciones de las extremidades anteriores para la escalada que tenía Australopithecus. Su expansión mundial sugiere que Homo erectus era ecológicamente flexible para adaptarse y prosperar ampliamente en diferentes entornos. Como era de esperar, con Homo erectus se da un importante aumento en el tamaño del cerebro, hasta 1250 centímetros cúbicos en los especímenes asiáticos. El tamaño de los molares se reduce en Homo erectus respecto a Australopithecus, que refleja su dieta más suave.
Hará unos 700.000 años, o quizás antes, Homo erectus en África dio lugar a Homo heidelbergensis, una especie muy parecida a la nuestra en términos de proporciones del cuerpo, adaptaciones dentales, y capacidad cognitiva. El Homo heidelbergensis, conocido como una especie arcaica de Homo sapiens, era un activo cazador de caza mayor, produce sofisticadas herramientas de estilo de Levallois, y por lo menos hace 400.000 años ya había aprendido a controlar el fuego. Los neandertales (Homo neanderthalensis) son homínidos adaptados al frío, con físicos fuertes, comportamientos complejos, y el cerebro de tamaño similar al nuestro, que ha evolucionado a partir de poblaciones europeas de Homo heidelbergensis, hará unos 250.000 años.
Los fósiles y las pruebas de ADN indican que nuestra propia especie, Homo sapiens, evolucionó en África hace 200.000 años, probablemente de Homo heidelbergensis. El aumento de la sofisticación del comportamiento de Homo sapiens, con cerebros grandes (1.400 cc) y la evidencia arqueológica de un conjunto más amplio de herramientas y técnicas de caza inteligente, permite a nuestra especie a florecer y crecer en el continente africano. Hace 100.000 años, nuestra especie se expande por Eurasia, y con el tiempo a todo el mundo, colonizando Australia y las Américas. En el camino, nuestra especie se encuentra con otros homínidos que se encontraron, como los neandertales en Europa y formas similares en Asia. Pero de este proceso de interacción existen numerosas dudas. Los estudios de ADN extraído de fósiles neandertales sugieren que nuestra especie puede ocasionalmente haberse cruzado con ellos. Nuestro impacto global creciente continúa en la actualidad, como las innovaciones culturales tales como la agricultura y la forma de urbanización del paisaje y de las especies que nos rodean.
En resumen, la evolución de la especie humano desde el Mioceno fue un proceso complejo. Nuestro linaje está lleno de ramas laterales y callejones sin salida evolutivos, con especies como los australopitecinos robustos que persistieron por más de un millón de años antes de desaparecer. Algunos rasgos humanos, como el bipedismo, evolucionaron muy temprano, mientras que otros, como poseer grandes cerebros, no evolucionaron hasta hace relativamente poco. Sin embargo, otros rasgos, como el tamaño de los molares, han evolucionado en una dirección para cambiar de nuevo más tarde mediante el cambio de presiones ecológicas. En lugar de tener trazado un rumbo directo hacia un destino predeterminado, la evolución de nuestro linaje (de hecho, del linaje de cualquier especie) es más próxima a la imagen de un bote salvavidas sacudido por los mares cambiantes de los cambios ambientales, el azar de la genética y la oportunidad de un determinado período de la historia de la Tierra.
Hasta hace poco, los eventos evolutivos que rodearon el origen del linaje homínido, que incluye a los humanos modernos y nuestros parientes fósiles, eran prácticamente desconocidos, y nuestra relación filogenética con los actuales monos africanos ha sido muy debatida. Los gorilas y los chimpancés han sido considerados estrechamente relacionados entre sí, debido a su alto grado de similitudes morfológicas y de comportamiento, tales como su modo compartido de la locomoción, la caminata de nudillos. Sin embargo, tras los estudios moleculares se ha hecho evidente que los chimpancés comparten un ancestro común más reciente con los seres humanos, y son por lo tanto más estrechamente relacionados con nosotros que con los gorilas. Se pensaba que las similitudes entre los actuales simios africanos de vida se habían heredado de un ancestro común de características primitivas, lo que implicaría que los primeros homínidos y nuestro último antepasado común compartido con los chimpancés tenían características similares en morfología y comportamiento a los simios africanos. Tras los descubrimientos de especies de hominidos más tempranas en la historia evolutiva, estos supuestos están sometidos a crítica.
La fecha de divergencia chimpancé-humano se ha estimado que pudo ser hace entre 8 y 5 millones de años desde la década de 1960 a través de técnicas inmunológicas y moleculares. Debido a nuevas hipótesis basadas en la genética, se han producido intensos esfuerzos por parte de diferentes equipos en las últimas dos décadas para encontrar y explorar sedimentos que registren este período de tiempo para el que prácticamente no se tenía ninguna evidencia fósil. El arduo trabajo y la perseverancia llevaron al descubrimiento de varios nuevos géneros y especies de los primeros homínidos cerca de las fechas estimadas de divergencia de los chimpancés y los seres humanos. En 1994 fue descubierto el Ardipithecus ramidus, de hace 4,4 millones de años, y poco después, se descubrieron homínidos aún más antiguos: Orrorin tugenensis (5,7-6,0 millones de años), Sahelanthropus tchadensis (6-7 millones de años), y Ardipithecus kadabba (5,2 a 5,8 millones de años).
Estos primeros homínidos carecen de características derivadas que se encuentran en los homínidos posteriores, y su inclusión en el linaje homínido se basa en gran medida en la reducción de tamaño de los caninos y la presencia de adaptaciones morfológicas para la bipedestación habitual, en particular en la pelvis y extremidades posteriores. El bipedismo es a menudo considerado como el sello distintivo de los homínidos, y su presencia en especies fósiles es a menudo la clave para su inclusión en el clado homínido. Sin embargo, la condición de homínidos es cuestionada por diversos investigadores en el campo.
Sahelanthropus tchadensis
Descubierto por la Misión Paleoantropológica Franco-Chadiense, en el año 2001, en el sitio de Toros-Menalla, en Chad, puede ser el homínido más antiguo recuperado hasta el momento. Sin embargo, la edad de Sahelanthropus se determinó biocronológicamente. A diferencia de los sitios de África oriental, Toros-Menalla carece de tobas volcánicas, lo que excluye el uso de la datación radiométrica. Aunque se estima un rango de edad de 6-7 millones de años para Sahelanthropus tchadensis, los sedimentos en los que se hallaron los fósiles están en el extremo anterior de ese rango, de 6,8 a 7,2 millones de años.
Único cráneo existente de Sahelanthropus tchadensis
El único espécimen de de Sahelanthropus tchadensis, apodado Toumaï, tiene características que le hacen estar más cerca de los humanos que de los simios. Se cree que vivió en zonas pantanosas. Su cráneo es simiesco y pequeño, aunque de rostro pequeño y dientes pequeños, en particular los caninos, lo que le acerca a los humanos. Hay ausencia de espacio entre los dientes. Su arco supraorbital es muy prominente. Se ha encontrado un cráneo, dos fragmentos de mandíbula inferior y tres dientes aislados. No presenta cresta craneal, su volumen cerebral es aproximadamente de 350 cm³, similar a los chimpancés modernos y mucho menor al de los humanos (1.350 cm³). Tiene rostro alto y poco prognatismo. La inferida posición del cráneo con referencia al cuerpo seria un indicador de posición bípeda. Podría ser el antecesor de Ardipithecus ramidus.
En todo caso, los restos fósiles son escasos y para muchos especialistas, esos rasgos no tienen porque incluirle necesariamente en los homínidos, ni suponer que se acercaría a la bipedestación, sin poder valorar otros elementos anatómicos, como la pelvis y el fémur, ya que el valor taxonómico y la importancia funcional de la base del cráneo acortado y la posición del foramen magnum no están claros.
Dibujo del posible aspecto de Sahelanthropus tchadensis
Orrorin tugenensis
Los especímenes de Orrorin tugenensis fueron descubiertos en la Formación Lukeino de las colinas Tugen, Kenia por la Expedición Paleontología Kenya. Los sedimentos tenían una antiguedad de 5,7-6 millones de años. Entre ellos se encontraban fragmentos mandibulares, dientes aislados y algunos elementos postcraneales. En términos generales, la dentición es morfológicamente más similar a la de los simios, particularmente los caninos, ya que son grandes, triangulares y agudos.
Descripción de los fósiles de Orrorin tunegensis
Su posible bipedestación es objeto de debate a partir de sus elementos, concretamente un fémur proximal. Comparte varias características con los homínidos posteriores que se interpretan como indicadores de bipedestación: cuello alargado, presencia de tuberosidad glútea. En cuanto al fémur, indicaría una cierta bipedestación, sin embargo, muchos investigadores cuestionan la metodología empleada y la calidad de la tomografía computarizada utilizada.
Para algunos investigadores, los fémures son más parecidos a los humanos que los de Australopithecus, argumentando que Orrorin condujo directamente a Homo y, por lo tanto, relegando a Australopithecus a un lado de la rama del árbol evolutivo humano.
Recientes estudios morfométricos de Richmond y Jungers (2008) sugieren que Orrorin es morfológicamente similar a los homínidos del Plioceno, distintos de Homo y los grandes simios actuales, lo que implica que puede haber poseído una forma de bipedalismo similar a los homínidos del Plioceno.
Ardipithecus kaddaba
De la especie Ardipithecus kaddaba se conoce su existencia a finales del Mioceno, por fósiles descubiertos en las localidades de la margen occidental de Middle Awash, Etiopía. Las muestras consisten en fragmentos mandibulares, dientes aislados y unos pocos elementos postcraneales recuperados. Se ha datado en 5,2-5,8 millones de años.
En principio, fueron catalogados originalmente como una subespecie de Ardipithecus ramidus, pero más tarde fueron otorgados a una especie propia. Entre los elementos postcraneales recuperados está una falange proximal del pie, cuyas características la relacionan claramente con el bipedalismo. En cuanto a la dentición, la de Ardipithecus kaddaba es algo más primitiva que la Ardipithecus ramidus. A través del fechado de los fósiles, se considera que Ardipithecus kadabba es cerca de un millón de años más antiguo que el Ardipthecus ramidus.
Descripción de los fósiles de Ardipithecus kaddaba
Paleoambiente de los homínidos tempranos
Desde el descubrimiento de Australopithecus africanus y la recuperación de la fauna asociada indicativa de hábitats abiertos, se ha postulado que el origen del bipedismo en el linaje homínido tendría sus raíces en la expansión de las sabanas expansión en el Plioceno. Descubrimientos posteriores de Australopithecus afarensis y fauna asociada parecen corroborar esta hipótesis. Sin embargo, los contextos ecológicos de los primeros homínidos sugieren que la relación entre la aparición del bipedismo y la expansión de la sabana y de los hábitats de pastizales puede ser tenue.
Interpretaciones actuales del paleoambiente sugieren que Orrorin se asocia a hábitats forestales abiertos con densos bosques o bosques en las cercanías, posiblemente a lo largo de los márgenes del lago.
A Sahelanthropus es probable encontrarlo en un mosaico de ambientes, desde el bosque de galería en el borde de un lago, a la sabana arbolada, o pastizales abiertos, aunque hay indicios de que hubo un predominio de arbustos y matorrales y hábitats de bosques verdes dentro de la cuenca del lago Chad
Ardipithecus kadabba está asociado con el bosque de ribera y la llanura de inundación de pastizales a lo largo de márgenes de zonas con agua. Ardipithecus ramidus se encuentra en hábitats de bosques cerrados con posibles manchas de bosque en Aramis y se asocia con matorrales y los hábitats de pastizales en Gona.
Combinadas, estas interpretaciones paleoambientales de África, a finales del Mioceno y el Plioceno temprano sugieren que los comienzos de nuestro linaje no ocurrieron en zonas abiertas, con condiciones de hábitat semiáridas a áridas, sino más bien en ambientes cerrados y/o húmedos. Sin embargo, es difícil establecer una conclusión definitiva debido a la falta de reconstrucción paleoecológica detallada de Orrorin, la posibilidad de que Sahelanthropus fuera encontrado en hábitats más abiertos, la discordancia en la interpretación del conjunto de datos en Aramis, y la escasez general de lugares paleontológicos en el Mioceno tardío de África.
Mientras que la condición de homínido de las especies Sahelanthropus, Orrorin y Ardipithecus puede ser objeto de debate, su importancia es clara para la comprensión de los orígenes humanos y su evolución. Se abre una puerta hacia los orígenes de nuestro linaje, en el momento en que nuestros antepasados se separaron de nuestros primos los chimpancés. A medida que se recuperen y analicen más fósiles de estas especies y su fauna asociada, vamos a aprender más, no sólo sobre su paleobiología, taxonomía y relaciones filogenéticas, sino también sobre su entorno y la forma en que pueden haber influido en la evolución de nuestro linaje.
Homo sapiens es el único representante vivo de la tribu Hominini , pero esto no siempre fue así, nuestros antepasados se superponen en el tiempo y el espacio con otras especies en todo el Plio-Pleistoceno. Existen numerosos factores que pueden estar implicados en la evolución o extinción de una especie, pero el principal entre ellos son las condiciones ecológicas y la capacidad o incapacidad de una especie para adaptarse a los cambios. Es fundamental para nuestra comprensión de la evolución humana llegar a comprender la especiación, extinción, y la variabilidad morfológica y de comportamiento que se observa en el registro fósil y arqueológico.
Hay muchos enfoques para reconstruir los valores ecológicos y ambientales de la evolución humana y muchos de ellos se solapan en cierta medida. La ciencia de la paleoecología se ocupa de las relaciones entre especies de plantas y animales y sus ambientes, mientras que la investigación paleoambiental tiende a centrarse más en las condiciones físicas de los alrededores, incluyendo el terreno y la estructura de la vegetación, así como características tales como la temperatura y precipitaciones. La paleoclimatología, una disciplina relacionada, se refiere al clima del pasado y a su reconstrucción de amplios registros a largo plazo a nivel mundial o regional, pero también es importante investigar el efecto local de estas variables en los sitios donde se sabe que los homínidos han vivido.
Mamíferos: pasado y presente
Los mamíferos muestran las relaciones que existen entre las especies y sus entornos. Todas las especies de mamíferos requieren dos cosas importantes de su hábitat natural para la supervivencia diaria: recursos alimenticios para satisfacer sus necesidades de energía y espacio en que comer, dormir y moverse. Los dietéticas o trófica y espacial nichos que están disponibles para cada taxón que rellenar un lugar determinado se dictarán por bióticos factores tales como el tipo de vida vegetal y animal, presentes y abióticos factores tales como el tipo de suelo, la precipitación, la temperatura y la estacionalidad . Estos factores determinarán qué organismos pueden sobrevivir en un hábitat determinado.
Considerando dos tipos diferentes de ambientes africanos, un bosque y un bosque-matorral abierto, creados por las variaciones en los factores bióticos y abióticos, nos centraremos en los diferentes mamíferos que son capaces de vivir en cada uno de ellos y tomar ejemplos específicos de de ungulados, bóvidos y los carnívoros que se alimentan en ellos.
En latitudes tropicales donde la humedad y las altas temperaturas anuales crean condiciones de bosques con varios pisos de árboles y ciclos de fructificación y floración de la vegetación, habrá una gran cantidad de especies que están adaptadas para vivir en el sustrato arbóreo, y muchos de ellos serán frugívoros y arborícolas, alimentándose de frutas y hojas. Bóvidos pequeños como los antílopes están bien adaptados para estos ecosistemas forestales, por ejemplo, Cephalophus callipygus, consume principalmente frutas y sus depredadores naturales son especies de felinos de la familia Felidae, como el leopardo (Panthera pardus) y el gato dorado africano (Profelis aurata), estando ambos adaptados para moverse rápidamente por arriba y abajo de los árboles.
En cambio, cuando las condiciones son más áridas y la temperatura y las precipitaciones fluctúan estacionalmente, con condiciones más abiertas de arbolado y matorral, predominando el matorral y con existencia de pastos, la comunidad de mamífero tendría un mayor número de especies adaptadas para el pastoreo y la locomoción terrestre. Un bóvido se encuentra comúnmente en ecosistemas más abiertos es Nanger granti , gacela de Grant, que come las hierbas frescas cuando están estacionalmente disponibles y las hierbas y las hojas en los tiempos restantes del año. Esta especie es presa de felinos que acechan en las zonas abiertas, como el guepardo (Acinonyx jubatus) y el chacal (Canis aureus), que es un miembro de la familia de los perros, los cánidos. A pesar de que la gacela de Grant es un corredor rápido, no siempre puede evitar la depredación de estos carnívoros rápidos y sigilosos.
Observando el comportamiento alimentario y la forma de vida de varios taxones que habitan en una localidad, es posible evaluar la forma en que toda una comunidad de mamíferos ocupa nichos espaciales y tróficos, proporcionándonos una imagen de la estructura general de las comunidades de mamíferos. Los nichos tróficos y los nichos espaciales se subdividen en categorías indicativas de las adaptaciones dietéticas y locomotoras de cada especie y las proporciones de las distintas especies en cada categoría se calculan para facilitar la comparación entre las localidades.
Dentro de las adaptaciones del aparato locomotor, dentro de la locomoción terrestre, podemos observar la capacidad de escalada de los gatos o la adapatación para la excavación subterránea, para cavar madrigueras de algunos roedores. En el bosque tropical de Seredou, en Guinea, aproximadamente el 30% de las especies del bosque son terrestres, pero si se combina con la capacidad de movimiento en los árboles (arbórea, semi-arbórea y trepadora), hay el doble de especies (59%) que las específicamente terrestres. Adicionalmente, el bosque de Seredou posee proporcionalmente mayor cantidad de los frugívoros, que dependen de los árboles frutales. En contraste con Seredou, los mamíferos residentes en Banagi, sabana de Tanzania, son predominantemente terrestres (70%). Banagi también tiene un mayor número de especies herbívoras forrajeras y carnívoras, y dentro de estas últimas más abundantes los depredadores que cazan especies herbívoras forrajeras.
Para estudiar la paleoecología en el registro fósil, trabajamos hacia atrás en el tiempo mediante la evaluación de los recursos tróficos y adaptaciones del aparato locomotor de los mamíferos en una localidad determinada con el fin de reconstruir las condiciones ecológicas del pasado. Como los huesos y los dientes se suelen conservar como fósiles, basamos nuestra comprensión en como las formas generales y otras características medibles proporcionan información sobre la explotación de nichos específicos. Las morfologías esqueléticas y dentales muestran adaptaciones a nichos específicos y tipos de hábitats, son conocidos como ecomorfologías. Las ecomorfologías dentales nos hablan de la dieta que una especie que no sólo ha tenido adaptaciones en sus dientes, sino también su cráneo y aparato locomotor. Las ecomorfologías se evalúan tomando medidas directas y analizando estadísticamente estas con el fin de buscar correlaciones significativas con los aspectos de la ecología del organismo, tales como dietas especiales o locomoción en espacios específicos. A menudo los paleoecólogos se centran en la evaluación de las ecomorfologías mostradas por especies de un solo grupo taxonómico en conjunto, en lugar de un enfoque completo de toda la diversidad de la comunidad. Al seleccionar cuidadosamente una familia diversa, abundante y bien conservada en un yacimiento de fósiles, se evitan las dificultades que pueden afectar a los análisis basados en la comunidad, en particular, las condiciones tafonómicas, que a menudo eliminan ciertos tipos de especies con su información ecológica. Esto afecta comúnmente taxones de cuerpo pequeño, cuyos restos no son tan robustos como las especies más grandes y por lo tanto son a menudo destruidas de forma natural durante el proceso de fosilización. Algunos carnívoros también son capaces de destruir una parte importante de los restos identificables de sus presas, por lo que también pueden ser subrepresentadas en algunas circunstancias.
Laetoli
La reconstrucción de la diversidad de la comunidad ecológica y los enfoques ecomorfológicos son técnicas complementarias para la reconstrucción de las condiciones ecológicas en un lugar en un momento en el tiempo. Se debe estudiar la posición de los fósiles en las capas sedimentarias de forma que proporcionen información sobre la relación entre ellos.
Laetoli se encuentra al sur de la Garganta de Olduvai, en Tanzania. Es un lugar del período Plio-Pleistoceno y conserva las huellas y los restos de una comunidad diversa, con fauna que incluye los homínidos como el Australopithecus afarensis y Paranthropus aethiopicus en las capas sedimentarias superiores Laetolil (3,63-3,85 millones de años) y Ndolanya (2,66 millones de años).
Localización de la Garganta de Olduvai y Laetoli
El trabajo previo sobre las capas superiores Ndolanya ha identificado 44 especies de mamíferos en la comunidad. Estas especies se han catalogado según su peso corporal, patrón de locomoción y la dieta. se observa una alta proporción de mamíferos que se alimentan mediante el pastoreo. Las capas Ndolanya son únicas porque tienen un número mucho mayor de especies terrestres en comparación con los hábitats modernos, que pueden ser explicados por la pérdida tafonómica de algunas de las especies, probablemente las de cuerpo pequeño. Sin embargo, cuando se observa el patrón general de la estructura de la comunidad, se puede observar que las capas Ndolanya son similares a los hábitats áridos matorrales tropicales africanos. Estos también tienen una alta proporción de las especies terrestres de forma de vida arbórea, semi-arbórea y trepadora. Las capas Ndolanya representan un hábitat de matorrales áridos de hace 2,66 millones de años.
Las capas Laetolil, sin embargo, son aproximadamente un millón de años más recientes que las capas Ndolanya, y mucho puede cambiar en ese tiempo. Un análisis de la diversidad de la comunidad de estas capas, centrándose en los mamíferos más grandes de 500 gramos, que tienen más probabilidades de sobrevivir en el registro fósil, indica que las capas Laetolil tienen una mayor proporción de carnívoros que las capas Ndolanya pero que, en general, las capas son muy similares a las comunidades modernas del Serengeti en la forma en que los nichos tróficos y espaciales son explotados. En otros trabajos, sin embargo, se ha demostrado que la fauna Laetolil se asemeja más de cerca a las comunidades de los hábitats boscosos del Serengeti y no a los de las llanuras abiertas.
Las investigaciones ecomorfológicas de los bóvidos de Laetoli da peso a las conclusiones extraídas de los estudios de diversidad ecológica. Sin tener en cuenta la identidad de las especies presentes, las morfologías óseas de estos bóvidos indican que un mayor porcentaje de las capas superiores Laetolil, los bóvidos poseían adaptaciones para hábitats boscosos que los de las capas superiores de Ndolanya. Es interesante observar este cambio en los tipos de hábitat entre las dos capas durante un período de aproximadamente 300.000 años.
Muchos sitios paleontológicos poseen una secuencia cronológica claramente reconstruida y los fósiles se pueden asociar fácilmente con las unidades datables. Sus sedimentos y fósiles a menudo han sido depositados rápidamente a través de la acción del agua, tales como los meandros de los ríos o la expansión secuencial y la desecación de lagos antiguos. Sin embargo, Laetoli no se asoció con una fuente de agua permanente y los fósiles fueron depositados en la superficie y enterrados naturalmente. Aunque estas unidades de tiempo son bastante largas en comparación con otras secuencias geológicas, se hace evidente que la ecología no era invariable durante las capas Laetolil. Los matorrales son el grupo mejor representado de ecomorfologías, mientras que los bosques y las praderas no están bien representados. El cambio que se produce se refiere a los hábitats forestales. La cobertura de pastos fue aumentando a expensas de la cobertura arbórea, cambiando así la disponibilidad de los nichos tróficos y espaciales a los que estos bóvidos se adaptan mejor. Las adaptaciones para una gran variedad de hábitats se producen tanto por las comunidades Ndolanya y Laetolil, haciendo hincapié en que los ambientes no son homogéneos, pero en general se caracterizan por un mosaico de diferentes tipos de hábitat.
La región de Laetoli siempre se ha caracterizado por la variabilidad del hábitat, pero tuvo más cobertura boscosa a disposición de sus habitantes en el pasado que en la actualidad. La transición a un mosaico de hábitats más abiertos comenzó en el pasado y se observa en el registro fósil, tanto entre Laetolil Superior y Ndolanya, así como dentro de la secuencia superior de las capas Laetolil.
Los homínidos en Laetoli al principio probablemente se limitaron a una dieta vegetariana antes de adoptar una dieta basada en parte en los recursos animales cuando entraron en competencia con otros carnívoros. Además, un cambio en la cobertura arbórea, utilizada por muchos mamíferos no sólo para obtener alimentos básicos, sino como un lugar para dormir y buscar refugio, habría sido una modificación de los nichos espaciales que habría afectado a los ancestros de la especie humana. Adapatándose a las presiones selectivas de un componente de reducción del arbolado en sus ambientes, los homínidos evolucionaron a una mayor dependencia de los recursos alimentarios terrestres y una mayor eficiencia en la locomoción erguida.
Los análisis de los muchos sitios de África del período Plio-Pleistoceno sugieren que el Australopithecus afarensis vivió en bosques y matorrales hábitats parecidos a los de las capas Laetolil y que los Paranthropus habitaban en el extremo más abierto de estos ecosistemas, como vemos en Ndolanya. Los datos paleoclimáticos indican una tendencia a gran escala hacia una mayor variabilidad de hábitat, comenzando la aridez hace aproximadamente 2,8 millones de años. Los mamíferos de las capas Ndolanya pueden representar una comunidad que se habría adaptado a los cambios resultantes en los nichos ecológicos disponibles.
El dimorfismo sexual en las especies primates se define como las diferencias de tamaño existentes entre los individuos macho y los individuos hembra. Las diferencias de tamaño entre los sexos, además de sus implicaciones físicas, tienen implicaciones sobre la estructura y la organización social, rasgos evolutivos que no dejan huella en el registro fósil. Para determinar el grado de dimorfismo sexual es preciso contar con restos de, como mínimo, dos individuos y, además, que sean de distinto género, pues el grado de diferencia de tamaño entre sexos se mide a partir de la presencia de restos de individuos machos e individuos hembras. Se habla de especie monomórfica cuando las diferencias de tamaño son similares o pequeñas y de especie dimórfica cuando los tamaños son muy dispares.
Tradicionalmente, la norma general es que el macho sea de mayor tamaño que la hembra. De esta manera, los huesos más grandes corresponderían a los machos y los de menores dimensiones serían los de las hembras. Pero esta forma de operar sólo es válida para las especies con un marcado dimorfismo sexual. En especies poco dimórficas esto es más difícil de aplicar. Una vez determinado el sexo, para establecer el grado o media de diferenciación, nos podemos encontrar restos de individuos femeninos más grandes que los de individuos masculinos. En una especie poco dimórfica, por ejemplo la humana, las hembras grandes son mayores que los machos pequeños, mientras que en las especies muy dimórficas la más grande de las hembras no es mayor que el más pequeño de los machos.
El tamaño tiene implicaciones sobre la organización y distribución en el territorio de las diferentes especies de primates, estando la poligamia y la monogamia están relacionadas con el grado de dimorfismo. Los gibones, especie asiática de primates, presentan un grado de dimorfismo nulo, son monógamos y defienden juntos un territorio en el que solo admiten de parientes a sus crías. El gorila, homínido africano, es una especie muy dimórfica, el macho vive con varias hembras en un mismo territorio. El grado de dimorfismo sexual está directamente relacionado con la competencia de los machos por las hembras, de este modo los machos, con grandes dientes caninos, luchan frecuentemente por las hembras y son mayores que éstas. Se han realizado estudios sobre el grado de conflictos que se establecen entre las diferentes especies de antropoides en relación con el tamaño del diente canino, ya que es la pieza dental que mejor muestra las diferencias de tamaño.
Medir el grado de dimorfismo sexual en los homínidos no está exento de problemas. Incluso las especies con mejores registros fósiles, como Australopithecus afarensis y neandertales, presentan problemas. El tema está lejos de ser zanjado. La mayoría de los investigadores sostienen que el dimorfismo sexual de los australopitecinos era considerablemente mayor que lo que se observa en los seres humanos en la actualidad, pero otros investigadores argumentan que los australopitecinos muestran un nivel de dimorfismo bastante similar al de los humanos. La idea general es que, desde el desarrollo de las formas homínidas, a partir de Ardipithecus ramidus, hace 4,5 millones de años, el dimorfismo sexual ha descendido. Lo que no se sabe exactamente es si esa reducción de grado se ha dado de forma paulatina u ha ocurrido en algún punto preciso del árbol filogenético.
1. Ecuación de estimación de la masa corporal
La definición de dimorfismo sexual más utilizada es la de masa corporal, que viene a ser las diferencias, en promedio, entre hombres y mujeres en términos de tamaño total del cuerpo. El problema es que la masa corporal no está directamente preservada en el registro fósil.
Sin embargo, los elementos esqueléticos individuales, proporcionan datos para estimar la masa corporal. La cabeza del fémur, por ejemplo, es excelente para estimar la masa total del cuerpo. Esto no debería ser sorprendente, dado que un 70% de la masa total se apoya en la cabeza del fémur cada vez que se da un paso y se apoya el peso sobre un pie.
La siguiente ecuación se utiliza para estimar la masa corporal (BM) sobre la base de la amplitud de la cabeza femoral (FH, medida como la amplitud anterior-posterior) (tomado de McHenry, 1992):
BM (kg) = 2,239 × FH - 39,9
Trate de estimar la masa corporal en el modelo siguiente (sin clasificar) antes de pasar a las siguientes preguntas:
Espécimen 1 - Amplitud de la cabeza del fémur = 51,4 mm
¿Cuál es la estimación de la masa corporal para esta muestra?
2. Estimación de la masa corporal para varias muestras
Estimar la masa corporal sobre la base de las dimensiones de la cabeza femoral para los siguientes ocho muestras:
- Muestra 1: FH = 46,1 mm
- Muestra 2: FH = 39,8 mm
- Muestra 3: FH = 38,7 mm
- Muestra 4: FH = 53,5 mm
- Muestra 5: FH = 44,9 mm
- Muestra 6: FH = 50,1 mm
- Muestra 7: FH = 48,1 mm
- Muestra 8: FH = 49,9 mm
3. Dimorfismo sexual
Suponiendo que de las muestras anteriores, las muestras 1, 3, 5 y 6 corresponden a hembras, y las muestras 2, 4, 7, y 8 corresponden a machos, ¿cuál es el rango (o máxima variación) observable aproximado de las masa de los cuerpos de machos y hembras, en kg? ¿cuál será la masa corporal estimada promedio para machos y hembras?
Las estimaciones de masa corporal no hay que considerarlas como cifras exactas, ya que tienen cierto grado de error o incertidumbre. Vamos a suponer que el error promedio o incertidumbre en la estimación anterior sería de +/- 4%. Por ejemplo, si se estima que un especimen tiene una masa corporal de 100 kg, sería realmente una estimación de 96-104 kg.
Si utilizamos las estimaciones mínimas (es decir, la estimación calculada, menos del 4%) de la masa corporal para nuestros individuos del sexo masculino identificados, ¿cuál es la masa corporal media estimada masculina? ¿la femenina? ¿la promedio de las dos?
4. Registros fósiles
Una técnica utilizada es la aplicación de la técnica del "bootstraping" o aleatorización muestral. Por medio de ella se sacan, de forma aleatoria, un número alto de parejas (triadas si la muestra fósil es de tres individuos, etc.) cuyas medidas están registradas en una base de datos y comprobar en cuantas parejas, triadas, etc. se produce un grado de variación como el de las muestras halladas en el yacimiento. Para ello, las medidas han de tomarse de muchas y diversas partes del cuerpo porque éste no representa una medida única.
Se puede contrastar la variación de los datos comparándolos con los de especies dimórficas, como los gorilas o monomórficas, como la humana. Así, la variación de Australopithecus afarensis sería similar a la de los gorilas, por lo que se podría suponer que los rasgos sociobiológicos de los gorilas se asemejarían a los de los Australopithecus afarensis. Por el contrario, comparando otras especies con la humana, se supondría que el inicio de la reducción del dimorfismo sexual se habría dado con Homo ergaster (hace 1,8 millones de años), reduciéndose progresivamente y alcanzándose ya con Homo heidelbergensis (hace 300.000 años) valores próximos a los humanos.
El dimorfismo no tiene equivalencia en todos los elementos esqueléticos. Algunos elementos esqueléticos permiten mayor facilidad para la identificación de la anatomía masculina y femenina. La capacidad de distinguir machos y hembras es muy importante a la hora de investigar el dimorfismo.
Tenemos dos muestras a partir de moldes de las cabezas de los fémures de dos especimenes típicos de Australopithecus afarensis. La amplitud de la cabeza femoral de las dos muestras es la siguiente:
- AL 288-1 ("Lucy") = 28,0 mm
- AL 333-3 = 41,0 mm
Estímense las masas corporales de los dos especímenes.
El AL 288-1 ("Lucy") corresponde a una hembra y el AL 333-3. Compárense la variación de sus valores con los de la especie humana.
Hay que tener en cuenta que la muestra fósil disponible es mayor que sólo estos dos ejemplares, y estos dos podrían representar una imagen un tanto exagerada de la variación típica en Australopithecus afarensis. Sin embargo, en una muestra más grande, la incertidumbre de las mediciones individuales, así como la incertidumbre de las clasificaciones de macho y hembra se convierten en un problema más grande. Todo ello significa que el dimorfismo es difícil de calcular con precisión, aunque la mayoría de las estimaciones sugieren que el dimorfismo en Australophitecus afarensis excede al observado en los seres humanos.
Si tenemos en cuenta que los machos de gorila pesan entre 140 y 200 kg y las hembras la mitad; en Australophitecus afarensis el promedio de masa corporal era, para machos 45 kg y para hembras 29 kg; comparénse los datos.
Se estima un umbral de 5% de diferencia entre machos y hembras en especies que no son dimórficas, como la humana. ¿Ocurre esto es Australopithecus afarensis?
Como conclusión, el dimorfismo sexual de Australopithecus afarensis, está más próximo al de los gorilas o al de los humanos?
Ardipithecus ramidus es una especie extinta de homínido, probablemente un hominino (primate bípedo) y quizá un ancestro del hombre. "Ardi" significa suelo, "pithecus" en griego significa mono y ramid es raíz, en la lengua (amhárico) del lugar (Etiopía) donde fueron encontrados los primeros restos.
Ardipithecus ramidus puede ser una especie descendiente de Ardipithecus kadabba. Sus restos se encontraron en la localidad de Aramis (con una antigüedad de 4,4 millones de años) en Middle Awash, Etiopía, y en la localidad de As Duma (con una antiguedad de 4,32-4,51 millones de años) en Gona, Etiopía.
Los principales rasgos que unen Ardiphitecus ramidus con los homínidos posteriores incluyen: caninos pequeños contundentes, dimorfismo reducido en cuanto al tamaño de los caninos, y caracteres indicativos de bipedestación. Pero Ardipithecus ramidus también presenta una serie de caracteres primitivos, sobre todo en el postcráneo, dedo gordo del pie plenamente oponible, ausencia del arco longitudinal del pie, similar longitud de las extremidades posteriores y una pelvis simiesca. Por lo tanto, muestra un mosaico de rasgos avanzados y primitivos que sugieren que era un bípedo facultativo, capaz de trepar en los árboles. Esto refuta las suposiciones anteriores de que el último ancestro común de chimpancés fue similar, y en cambio sugiere que los chimpancés son organismos altamente especializados.
Se ha observado, sin embargo, que Oreopithecus bambolii, mono de finales del Mioceno, cuyos restos se han encontrado en Italia, muestra muchos de los caracteres que se utilizan para indicar el estado de homínido de Ardipithecus ramidus, como relativamente pequeños caninos, colocación del foramen magnum más próxima a los homínidos y un buen desarrollo de la espina ilíaca anterior inferior. Mientras que otras características morfológicas de Oreopithecus no dejan ninguna duda de que es un mono, la presencia de los caracteres anteriores sugiere que las interpretaciones de los primeros homínidos pueden ser confusas.
Cráneo de Ardipithecus ramidus
Por análisis de la composición isotópica del suelo, muestras y dientes, Ardiphitecus ramidus era un habitante de bosques con pequeños claros. Su dieta era probablemente más omnívora que la de los chimpancés (especialistas en fruta madura) y es probable que se alimentara tanto en los árboles como en el suelo. Aparentemente consumía pequeñas cantidades de recursos en medio abierto, con el argumento en contra de la idea de que un poblamiento de las praderas fue la causa del origen de caminar erguido.
Ardipithecus ramidus, descrito por primera vez en 1994 a partir de los dientes y fragmentos de una mandíbula, hoy está representado por 110 especímenes, incluyendo el esqueleto parcial de una hembra, rescatado de la degradación por erosión. Este individuo pesaba unos 50 kg y medía unos 120 centímetros de alto. En el contexto de los muchos otros individuos recuperados de esta especie, esto sugiere un cuerpo pequeño, con diferencia de tamaño entre machos y hembras. El tamaño del cerebro era tan pequeño como en los chimpancés actuales. Los numerosos dientes recuperados y una gran parte cráneo completo muestran que Ardipithecus tenía una cara pequeña y un menor tamaño de canino, con un premolar complejo, indicativo de agresión social mínima.
Sus manos, brazos, pies, la pelvis y las piernas revelan que se movía hábilmente en los árboles, con el apoyo de sus pies y palmas de las manos. Era palmigrado y escalaba, pero carecía de las características típicas de la suspensión, escalada vertical, o caminata de nudillos de los gorilas modernos y chimpancés. Su hábitat fundamental era terrestre, con una forma de bipedalismo más primitivo que el de los australopitecos. Ardipithecus ramidus indica, pues, que el ancestro común de los humanos y los simios africanos no era similar a un chimpancé, ya que tanto los homínidos y los simios africanos existentes son cada uno muy especializados, pero a través de muy diversas vías de evolución .
Un punto esencial para el estudio de la posición erguida o erecta son los músculos glúteos, siendo función del músculo glúteo mayor la extensión de la pierna detrás del cuerpo y mantener el cuerpo erguido, y la del menor, el equilibrio durante la marcha.
Músculos glúteos mayor y medio
Si se observa el extremo superior de un fémur, se podrá distinguir, entre otros elementos, la cabeza articular, esférica y lisa, el trocánter mayor, saliente óseo cuboideo, muy aplanada transversalmente, situada por fuera y un poco por debajo de la cabeza articular, su cara externa, convexa, presenta la cresta del glúteo medio, dónde se inserta este músculo.
El glúteo mayor se inserta en la línea áspera, borde del fémur, grueso, saliente y rugoso, que separa perfectamente la cara externa de la cara interna.
Fémur humano
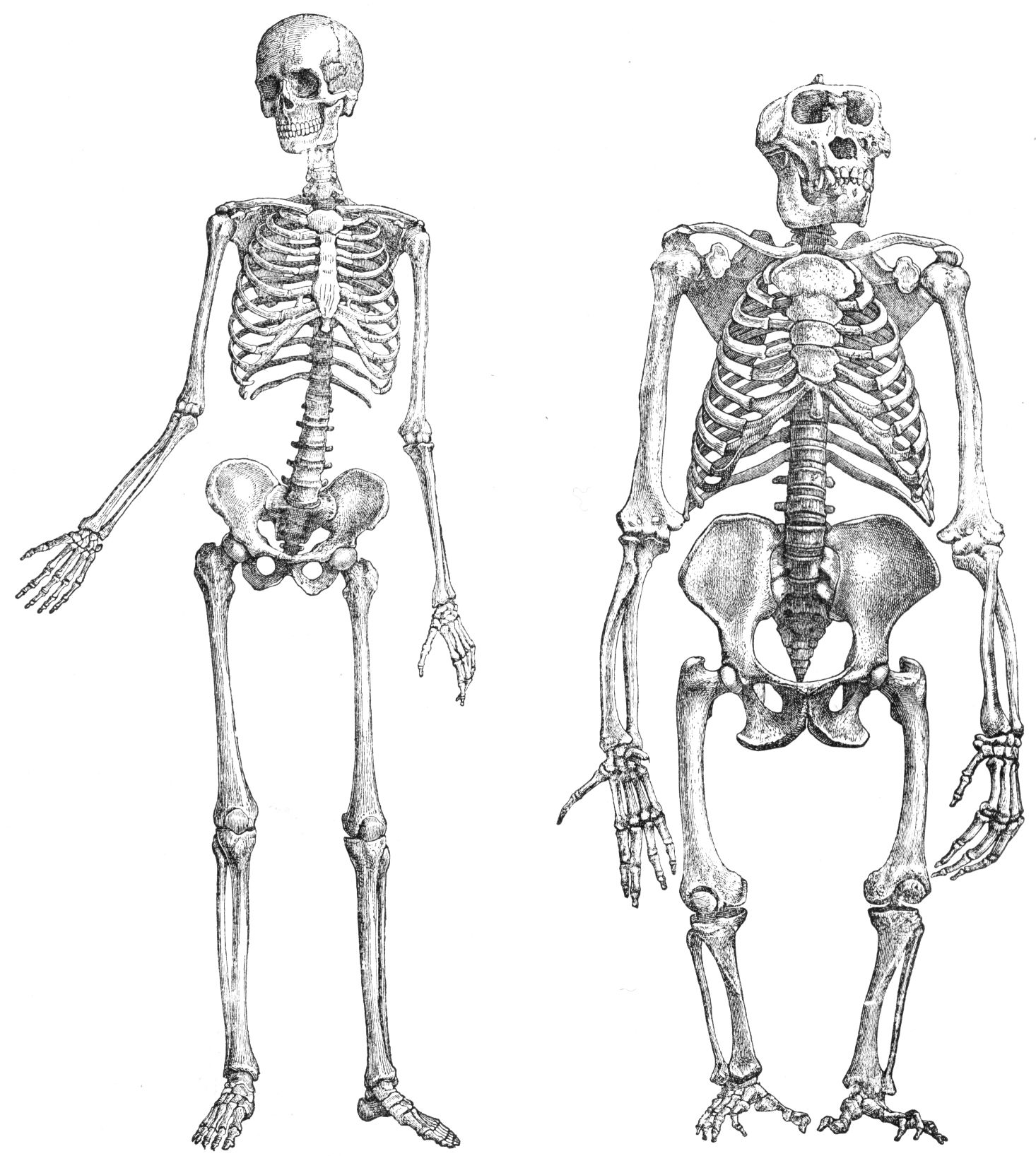
Comparando los esqueletos humano (izquierda) y del gorila (derecha) ponga de relieve las diferencias en cuanto al fémur y a la inserción de los músculos glúteos en el mismo.
2. Pelvis
Otro cambio importante, asociado a la bipedestación y a la postura erecta aparece en la pelvis. Ésta se ensancha para soportar mejor el peso de los órganos al estar en posición erecta. Al mismo tiempo, su posición de modifica, por lo que las crías nacen prematuras. En los otros mamíferos el llamado canal de parto es muy breve, y a medida que la pelvis va evolucionando, este se hace prolongado y sinuoso, lo que hace dificultosos los alumbramientos.
Pelvis de Australopithecus sediba
Esqueleto de orangután
Comparando el esqueleto de orangután y la pelvis de Australopithecus sediba, poner de relieve las diferencias existentes en cuanto a la pelvis y mostrar su evolución.
3. Estructura del pie
El otro suceso iniciador en el movimiento hacia delante de la extremidad posterior es el "despegue de los dedos". Como se inicia una zancada bípeda, antes de abandonar el suelo, el pie proporciona una considerable fuerza de propulsión a través del margen anterior del pie, y en particular el dedo gordo del pie (o hallux). En los seres humanos, este despegue de los dedos se logra a través de un pie bastante rígido que transmite la fuerza efectiva desde el talón hasta el dedo del pie. Los chimpancés, que utilizan sus pies en la escalada, caminata de nudillos, y la bipedestación, tienen una estructura del pie mucho más generalizada.
Observando los anteriores esqueletos de gorila y humano, poner de relieve las diferencias anatómicas de sus pies.
4. Hueso calcáneo
El hueso calcáneo es un hueso del pie (tarso), corto, asimétrico, de forma cúbica irregular, con seis caras: superior e inferior, laterales y anterior y posterior, de las cuales dos son más o menos articulares. Este hueso constituye el talón del pie.
Dada la mayor transferencia de peso y la fuerza involucrada en el golpe de talón bípedo, se da una adaptación biomecánica en los homínidos bípedos. La mayor fuerza sobre un talón bípeda requiere una superficie más amplia para el talón, con el fin de distribuir más eficazmente esta fuerza.
Observando los anteriores esqueletos de gorila y humano, poner de relieve las diferencias anatómicas en cuanto a sus huesos calcáneos.
5. Huellas
Las huellas reflejan la estructura anatómica del pie y por tanto nos permiten hacernos una idea sobre el grado de bidepestación de una especie.
Las líneas de huellas de homínidos de Laetoli fueron descubiertas en 1976-1977 por Mary Leakey, Richard Hay y su equipo. Fueron preservadas en ceniza por una erupción del volcán Sadiman, a 20 km de allí. Una suave lluvia convirtió una capa de 15 cm de grueso en toba calcárea sin destruir las impresiones. Con el transcurso del tiempo fueron cubiertas por otros depósitos de cenizas.
Las huellas homínidas fueron producidas por tres individuos, uno caminando sobre las huellas de otro, haciendo que la pistas originales fueran difíciles de encontrar
Huellas de Australophitecus en Laetoli
Observando las huellas, en cuanto al golpe de talón, ¿serían indicadoras de bipedestación?
CUVSI te ofrece 31 cursos online y gratuitos, con materiales libres, con certificados e insignias digitales, y no sólo eso, sino que quiere que estos adquieran valor real con la posibilidad de dar a conocer tus trabajos.
Necesitamos todos los apoyos para que este proyecto pueda crecer, para que sea una realidad sin fronteras, para crear un mundo formación, de saber y de investigación, un mundo mejor. Por ello necesitamos todos los apoyos para poder seguir ofreciendo cursos completamente gratis.
No te pedimos dinero, tan sólo que nos visites, o nos compartas en las redes sociales, cuantos más seamos mejor.
Gracias de todo corazón, por ayudar a crecer la educación con materiales libres en Internet y ayudar a divulgar el mundo del saber.
Cursos online, gratis, abiertos, con certificado e insignia digital